4.3 Redox Conditions
Microbially mediated oxidation-reduction reactions play a major role in regulating redox conditions in groundwater systems (Kuma and Riyazuddin, 2012; Liebensteiner et al., 2014; Tesoriero et al., 2015; Enright and Ferris, 2016; Enright et al., 2019). This is because microorganisms rely on the transfer of electrons from reduced electron donors to oxidized terminal electron acceptors as a source of energy for cell growth and division. The overall Gibbs energy of such reactions is determined by the difference between oxidant (electron acceptor) and reductant (electron donor) half-cell redox potentials (Equation 16). Those reactions that provide the maximum amount of energy are favored by microorganisms and typically dominate over competing reactions. The most common electron donors in groundwater systems are dissolved and particulate organic carbon (Shen et al., 2015). In some cases, reduced forms of nitrogen, iron, and sulfur are important. Among commonly available electron acceptors, oxygen yields more energy than any other oxidant in aerobic respiration. When oxygen is depleted, the next most energetically favorable electron acceptor in anaerobic respiration is nitrate, followed by manganese, ferric iron, sulfate, and finally carbon dioxide.
The sequential nature of electron acceptor utilization by microorganisms, which was introduced in Section 3 as the “redox ladder,” tends to promote the development of increasingly reducing conditions along groundwater flow paths depending on the availability of electron donors (Figure 13). As redox conditions change, documenting the sequential depletion of oxidants and corresponding production of distinctive conjugate reductants can be used to identify specific zones where microbial activity is dominated by a single electron-accepting process (Groffman and Crossey, 1999; McMahon and Chapelle, 2008). Characteristic threshold concentrations that have been suggested for the identification of redox zones in groundwater systems are given in Table 2. It is important to keep in mind that some care should be exercised when interpreting these threshold concentrations to account for factors such as differences in microbial community composition, availability of electron donors, and scale at which groundwater sampling occurs.

Figure 13 – Evolution of redox conditions along groundwater flow paths. Typically, redox conditions tend to become more reducing with increasing travel distance and residence time of water underground. Redox conditions in discharge areas tend to be variable owing to mixing of groundwater and surface water.
Table 2 – Threshold concentrations suggested for identifying microbial redox processes in groundwater systems (McMahon and Chapelle, 2008).
| Concentration mg/L | ||||||
| Redox Process | O2 | NO2− | Mn2+ | Fe2+ | SO42− | |
| Aerobic (oxic) | ||||||
| O2 Reduction | ≥ 0.5 | < 0.05 | < 0.1 | |||
| Anaerobic (anoxic) | ||||||
| NO3− reduction | < 0.5 | ≥ 2.2 | < 0.05 | < 0.1 | ||
| Mn(IV) reduction | < 0.5 | < 2.2 | ≥ 0.05 | < 0.1 | ||
| Fe(III)/SO4− reduction | < 0.5 | < 2.2 | ≥ 0.1 | ≥ 0.5 | ||
| CO2 reduction | < 0.5 | < 2.2 | > 0.1 | < 0.5 | ||
The first electron acceptor consumed along groundwater flow paths is oxygen. The transition from aerobic to anaerobic respiration is thought to take place when oxygen concentrations fall below a level of about 0.5 mg/L. For the onset of nitrate reduction (denitrification), the threshold oxygen concentration might be as high as 2.0 mg/L, implying a moderate degree of overlap with aerobic respiration. This is consistent with the growth of some heterotrophic microorganisms as facultative anaerobes, which utilize both aerobic and anaerobic respiration. The reduction of nitrate is also used by various lithotrophs in the oxidation of inorganic electron donors such as sulfide or ferrous iron. Sources of nitrate in groundwater include agricultural fertilizers, as well as lithotrophic microbial oxidation of ammonium released from the decomposition of nitrogen-rich organic substances by microorganisms.
In groundwater systems, Mn(IV) and Fe(III) are relatively abundant but typically occur as insoluble oxide mineral precipitates instead of dissolved ionic species. Because of this, elevated concentrations of dissolved Mn2+ and Fe2+ produced from the reductive dissolution of iron and manganese oxides are used as proxies to identify zones of active microbial Mn(IV) and Fe(III) reduction. The susceptibility of Mn2+ and Fe2+ to chemical and microbial oxidation, mineral precipitation, and solid-phase sorption processes add another layer of biogeochemical complexity that could result in an underestimation of the extent of Mn(IV) and Fe(III) reduction.
After oxygen and nitrate are exhausted, it is sometimes difficult to distinguish between zones of active Fe(III) and sulfate reduction. This relates to the energetic dependence of Fe(III) reduction on the crystalline form of iron oxides present in a groundwater system, as well as the natural variability in dissolved sulfate concentrations. As a general rule of thumb, poorly ordered hydrous ferric oxides are more easily reduced by microorganisms than crystalline varieties such as goethite or hematite (Roden et al., 2004; Langley et al., 2009a). These low-complexity ferric oxides can be thought of as more bioavailable, in the same way that simple monosaccharide sugars are more suited to yeast microorganisms than long-chain polysaccharides. Additionally, higher sulfate concentrations (≥ 0.5 mg/L) are energetically more favorable to sulfate reduction. Elevated concentrations of sulfide can also be used as a proxy for sulfate reduction, subject to the same caveats that apply to Mn2+ and Fe2+ proxies for Mn(IV) and Fe(III) reduction, respectively.
The redox conditions under which carbon dioxide reduction becomes the predominant anaerobic respiratory process are typically highly reducing owing to depleted concentrations of more strongly oxidizing electron acceptors such as sulfate (< 0.5 mg/L). It is tempting to use the presence of methane as a proxy for microbial carbon dioxide reduction; however, this can be misleading as some methanogenic microbes produce methane from the fermentation of acetic acid instead of the reduction of carbon dioxide. Other microorganisms produce acetic acid from carbon dioxide reduction instead of methane reduction.
An additional way to follow the shift in redox conditions along a groundwater flow path is to follow the change in microbial community composition from the unsaturated vadose zone to the saturated zone immediately below the water table. This can be accomplished using molecular biological techniques to identify and quantify different metabolic groups of microorganisms. A study on a shallow pristine sand aquifer found the relative abundance of microorganisms using oxygen as an electron acceptor decreases as water infiltrates through the vadose zone and becomes increasingly isolated from the atmosphere (Figure 14). The apparent preference for nitrate as an electron acceptor can be traced to the ability of some microorganisms to grow as facultative anaerobes. Below the water table, utilization of oxygen as an electron acceptor decreases while nitrite, Fe(III), and sulfate become more important. When it comes to electron donors (the energy source), organic matter and heterotrophic processes dominate above and below the water table. With the exception of Fe(II)-oxidizing microorganisms in the unsaturated zone, the relative abundance of lithotrophs using inorganic electron donors is lower than for their heterotrophic counterparts. Reduced Fe(II)-bearing minerals such as biotite and amphiboles are candidates as possible sources of Fe(II) to support lithotrophic microbial growth, particularly under relatively oxidizing conditions above the water table.
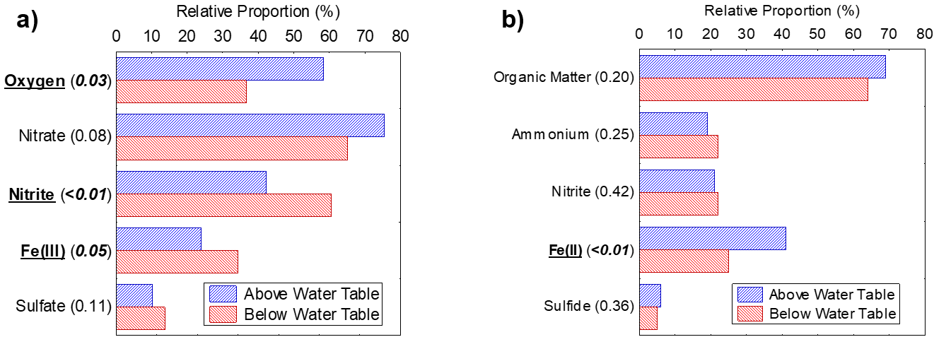
Figure 14 – Relative proportion of potential a) electron acceptor and b) electron donor use among bacteria in core samples obtained from above and below the water table in a shallow pristine sand aquifer on the Canadian Shield. The p-values of t-tests for the difference in average relative proportions of bacteria above and below the water table are given in brackets; significant values (p < 0.05) are italicized and corresponding metabolic groups are underlined in bold (adapted from Shirokova and Ferris, 2013).
Microbial contributions to redox transformations in groundwater systems are not strictly confined to the common electron acceptors that dominate in anaerobic respiration. In fact, possible oxidants for metabolic processes under reducing conditions include a wide variety of inorganic and organic substances, many of which are environmental pollutants. The conjugate reductants arising from the use of these atypical electron acceptors by microorganisms often display different physical, chemical, and biological properties than the parent compounds. Using chromium as an example, microbial reduction of Cr(VI) in the form of dissolved chromate (CrO42-) to Cr(III) produces the Cr3+ ion, which is highly insoluble. An important difference between the Cr(VI) and Cr(III) oxidation states is that hexavalent chromium is highly toxic, whereas trivalent chromium is an important trace nutrient. Arsenate (AsO43-) is another noteworthy electron acceptor in groundwaters because arsenic poisoning is a risk to human health (Gorra et al., 2012). Among organic contaminants in groundwater, highly oxidized chlorinated solvents such as tetrachloroethene (PCE) and trichloroethene (TCE) are known to behave as electron acceptors and undergo stepwise reductive dechlorination in microbial metabolism. While this contributes to the attenuation of PCE and TCE, an undesirable outcome in some settings is the production and accumulation of the carcinogenic intermediate, vinyl chloride (VC).